Physico-chimie dynamique de l’exocytose vésiculaire de neurotransmetteurs
Introduction
Introduction
L’exocytose vésiculaire joue un rôle central dans un grand nombre de phénomènes liés à la communication cellulaire au sein des êtres vivants complexes. C’est en particulier le cas dans les neurones (figure 1), dans les jonctions neuromusculaires ou dans la gestion de certains flux hormonaux. L’importance de ces flux de messagers chimiques est parfaitement reconnue par les biologistes et les médecins. Néanmoins, peu de chose est réellement connu sur les mécanismes exacts qui gouvernent les processus exocytotiques qui permettent aux cellules de délivrer ces messagers dans leur environnement extracellulaire.
Cette carence relative se comprend très bien dès que l’on prend conscience qu’une cellule ne délivre que de très faibles quantités de ces messagers : de quelques milliers (zeptomoles) à quelques millions (attomoles) de molécules seulement, ce qui représente une performance énergétique notable pour une cellule, mais une quantité insignifiante par rapport aux performances des meilleurs instruments analytiques. Le problème analytique est même encore plus complexe, puisque ces quantités sont délivrées en quelques millisecondes seulement et à un moment choisi par la cellule…
En fait le même problème se pose à la nature elle-même. En effet, dans la communication neuronale, dans les jonctions neuromusculaires, etc., les quantités délivrées par la cellule émettrice doivent être détectées par la cellule réceptrice afin de provoquer chez cette dernière la réponse adéquate. En biologie, on a tendance à » évacuer » cette difficulté en invoquant la très grande sélectivité des récepteurs dont est munie la cellule réceptrice. Bien sûr cela est vrai et cela seul conditionne la sélectivité de la reconnaissance : il suffit par exemple de se référer à ce brave papillon capable de détecter une seule cellule de phéromone de sa femelle grâce à l’extrême sélectivité des récepteurs dont sont munies ses antennes.
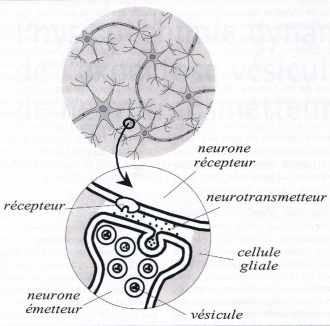
| Figure 1 |
| Représentation schématique d’une synapse neuronale. Le neurotransmetteur est contenu dans les vésicules présentes au niveau de la jonction terminale du neurone émetteur (en bas de la zone agrandie). L’arrivée d’un influx nerveux provoque l’ouverture de canaux ioniques dans la membrane du neurone, ce qui autorise l’entrée d’ions calcium dans la zone terminale du neurone. Ces derniers provoquent la fixation des vésicules à la membrane cellulaire puis la création d’un pore transmembranaire par lequel le neurotransmetteur est relâché dans l’espace synaptique où il diffuse et va activer un récepteur membranaire du neurone receveur, ce qui déclenche la propagation d’un nouvel influx nerveux dans le neurone récepteur (en haut de la zone agrandie). |
| Schéma 1 |
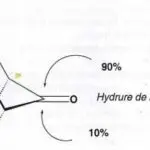
 |
| Principe de la détection électrochimique des molécules de catécholamines. L’oxydation globale à deux électrons de chaque molécule atteignant l’ultramicroélectrode placée au contact de la cellule émettrice en configuration de “ synapse artificielle ” (voir montage sur la figure 3, gauche) se traduit par un courant anodique (figure 3, droite ; figure 4) qui transcrit le flux d’émission cellulaire. |
Néanmoins, on oublie alors de rappeler que dans la nature ce fameux papillon est obligé de brasser des mètres cubes et des mètres cubes d’air avant de détecter par hasard cette molécule tant désirée. La vitesse avec laquelle le papillon trouverait l’objet de ses désirs serait nécessairement plus grande si cette molécule avait été délivrée dans un espace plus restreint. Cette remarque montre à l’évidence qu’à sélectivité et sensibilité du récepteur données, la détection est essentiellement liée à la concentration de l’espèce à détecter et non pas à sa quantité.
Dans la communication neuronale, dans les jonctions neuromusculaires, etc., la nature a résolu ce problème de cinétique en créant les synapses. En effet, qu’est-ce qu’une synapse ? Rien de plus qu’un nano-domaine confiné de quelques dizaines d’attolitres de volume, dans lequel le neurotransmetteur est délivré par la cellule émettrice. On conçoit alors que quelques milliers de molécules seulement suffisent à y créer un saut de concentration de l’ordre de 0,2 millimole par litre (0,2 mm), et partant plus que capable d’assurer une cinétique rapide.
C’est cette astuce retenue par l’évolution que nous avons empruntée. En effet, la cinétique électrochimique (c’est-à-dire l’intensité du courant) est uniquement fonction de la concentration de l’espèce détectée, et non pas de la quantité de cette espèce comme c’est par exemple le cas pour la plupart des autres méthodes analytiques et en particulier pour l’ensemble des méthodes spectroscopiques. Par conséquent, en plaçant une électrode de taille proportionnée à celle d’une cellule (c’est-à-dire généralement de quelques microns de diamètre, 7 µm pour les électrodes utilisées ici ; on parle alors d’ultramicroélectrode) au voisinage immédiat de cette cellule, on constitue une » synapse artificielle « . Pourvu que l’espèce délivrée par la cellule soit électroactive, l’ultramicroélectrode se comportera donc comme un neurone récepteur et pourra retranscrire fidèlement le flux d’émission de la cellule émettrice avec la précision et l’exactitude souhaitées.
Au Département de chimie de l’ENS, au sein de l’UMR » Pasteur « , nous utilisons cette méthode de » synapse artificielle » mise au point en collaboration avec notre collègue et ami Mark Wightman de l’université de Caroline du Nord à Chapel Hill (USA), dans le cadre de deux grands projets. L’un concerne la dynamique de l’exocytose vésiculaire de neurotransmetteurs, phénomène clef de la transmission synaptique ; l’autre concerne le stress oxydatif, phénomène naturel qui est entre autres à la base du pouvoir bactéricide des macrophages dans l’organisme, mais dont la disruption peut entraîner des troubles pathologiques graves (altération des codes génétiques, vieillissement, plusieurs types de cancers, les maladies de Parkinson ou d’Alzheimer, le sida, etc.).
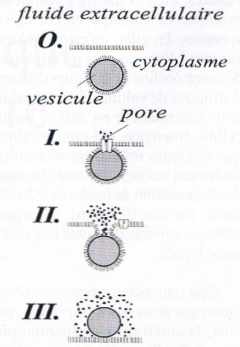
| Figure 2 |
 |
|
Représentation schématique des trois phases (I‑III) séquentielles d’une exocytose vésiculaire : (I) connexion initiale via un canal ionique ; (II) fusion des membranes cellulaire et vésiculaire ; (III) démasquage complet de la matrice polyélectrolyte de la vésicule et exposition totale au fluide extracellulaire. Les phases II et III correspondent approximativement à la montée rapide (II) puis à la descente lente (III) des flux d’émission mesurables via les variations de courant des figures 3 et 4. La phase I n’est pas observable sur les figures 3 et 4 car les échelles utilisées sont trop grandes par rapport à l’intensité (5 pA) et à la durée (1 à 2 ms) du flux d’émission pendant cette phase. |
On pourrait s’étonner a priori de voir de tels travaux » biologiques » abordés dans un Département de chimie et par des physico-chimistes. Cependant il s’agit là d’une démarche assez fréquente de nos jours où les frontières classiques interdisciplines n’ont plus grand sens. On ne peut plus aborder un problème transverse comme les deux thématiques prédentes par le biais d’une seule discipline.
Ainsi, de nos jours, le chimiste ne peut plus se définir comme étant uniquement » celui qui fait de la chimie « , mais plutôt comme celui » qui sait la chimie et la physico-chimie et sait en utiliser les concepts » pour apporter des points de vue originaux et des éclairages nouveaux dans des aventures intellectuelles relevant, par la nature du problème posé, d’autres disciplines comme la biologie par exemple. C’est précisément ce que nous essayons de faire en appliquant nos démarches et savoir-faire de chimistes et de physico-chimistes à ces deux problèmes fondamentaux de la biologie. Dans la suite, nous nous limiterons cependant à une présentation de nos travaux sur l’exocytose vésiculaire de neurotransmetteurs.
Dynamique de l’exocytose vésiculaire de neurotransmetteurs
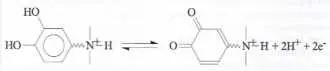
Dans les neurones, les neurotransmetteurs (catécholamines dans nos travaux ; voir structure moléculaire sur le schéma 1) sont » empaquetés » sous forme cationique dans des vésicules contenues à l’intérieur de la cellule émettrice (figure 1). Ces vésicules sont délimitées par une membrane analogue à celle de la cellule et contiennent une matrice polyélectrolyte anionique dans laquelle le neurotransmetteur cationique est piégé à haute concentration.
Nos travaux expérimentaux s’appuient essentiellement sur les cellules chromaffines des capsules surrénales plutôt que sur les neurones. Ces cellules qui sont impliquées dans la gestion des flux d’adrénaline dans l’organisme sont en effet considérées par les neurobiologistes comme de bons modèles pour l’étude de l’exocytose vésiculaire. Elles disposent de toutes les fonctionnalités décrites plus haut et sont plus facilement cultivables et isolables que les neurones. Dans ces cellules d’environ 10 µm de diamètre, les vésicules ont à peu près 150 nm de rayon et contiennent en moyenne 3 millions de molécules d’adrénaline.
La stimulation d’une cellule chromaffine par un flux d’ions divalent déclenche la création d’un pore transmembranaire nanométrique (1,5 nm de rayon selon nos résultats ; entre 1 et 2 nm de rayon sur la base de mesures indépendantes par patch-clamp) traversant les membranes cellulaire et vésiculaire au contact, et par lequel le neurotransmetteur est relâché dans le milieu extracellulaire (phase I sur la figure 2). Pour une raison inconnue jusqu’à nos travaux, ce pore peut devenir instable et sa rupture entraîner la fusion complète des deux membranes (phase II sur la figure 2).
Cette fusion se poursuit alors irréversiblement jusqu’à ce que la matrice polyélectrolyte soit complètement exposée à l’extérieur de la cellule sur la surface de sa membrane (phase III sur la figure 2). Ces trois phases avaient été mises en évidence auparavant grâce à des méthodes de mesure de capacité (dites de » patch-clamp »), puis plus récemment par spectroscopie fluorescente confocale à onde évanescente, mais la résolution cinétique de ces méthodes était insuffisante pour tenter d’observer la dynamique de la phase II, et encore plus d’identifier le phénomène physicochimique provoquant la transition de la phase I à la phase II.
Notre méthode de » synapse artificielle » (figure 3, gauche) permet de mesurer les flux de neurotransmetteurs émis par une cellule unique au cours d’une série de processus vésiculaires (figure 3, droite) et cela avec la précision cinétique voulue pour » démonter » finement le mécanisme de l’exocytose vésiculaire de neurotransmetteurs. Le flux des catécholamines relâchées est en effet détectable avec une résolution de l’ordre du millier de molécules par milliseconde grâce à leur oxydation électrochimique (schéma 1) à la surface d’une ultramicroélectrode placée en regard de la cellule émettrice. Le courant traversant l’ultramicroélectrode réceptrice reproduit alors fidèlement le flux d’émission vésiculaire (figures 3 et 4).
| Figure 3 |
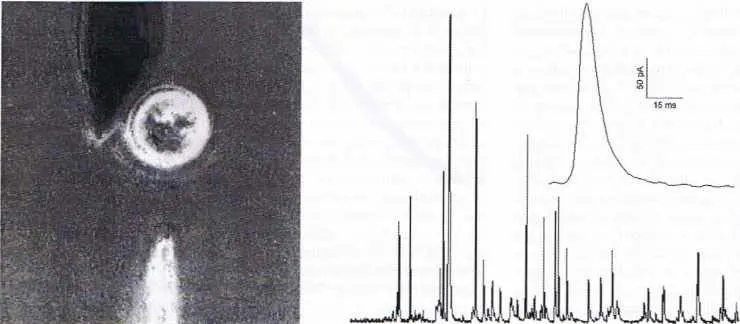 |
|
À gauche : synapse artificielle créée entre une cellule chromaffine (apparaissant comme un cercle clair sur la microphotographie) et une ultramicroélectrode (en noir, en haut de l’image) placée à son contact. La cellule est stimulée par une bouffée d’ions baryum réalisée injectée par une micropipette (en blanc, en bas de l’image). Cela déclenche une série d’émissions vésiculaires détectées ampérométriquement par l’ultramicroélectrode (schéma 1 ; traces courant (temps) représentées à droite de la figure). À droite : enregistrement continu de pics ampérométriques détectés par l’ultramicroélectrode. Chaque pic (voir détail agrandi de l’un d’entre eux sur la figure ; voir aussi la figure 4) correspond à un seul processus de vésiculaire. Échelle : la cellule chromaffine a un diamètre de 10 μm, et la surface active de l’électrode (partie placée en regard de la cellule et collectant le flux d’adrénaline) un diamètre moyen de 7 μm. |
| Figure 4 |
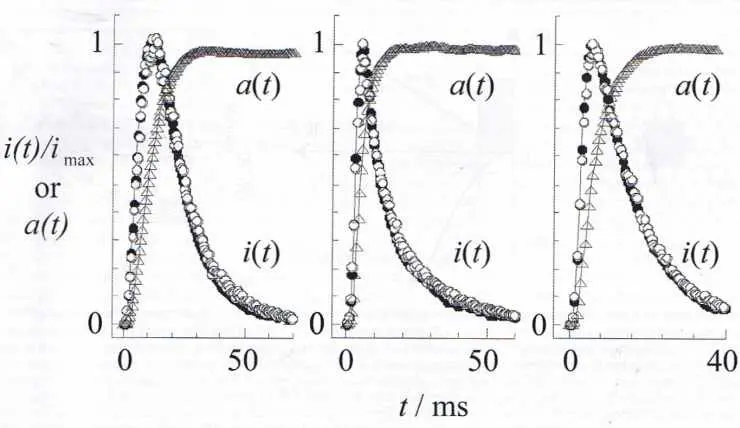 |
| Signaux ampérométriques mesurés sur une cellule chromaffine stimulée par Ba2+. Les courants expérimentaux (normalisés à leur maximum, i (t)/imax ; courbes en cloche) sont indiqués par les symboles circulaires, et le signal théorique calculé par la courbe pleine surimposée. Les courbes a (t) (allure sigmoïde, symboles triangulaires) représentent la variation de la fraction de la surface de membrane vésiculaire intégrée dans la membrane cellulaire à chaque instant (voir les figures 2 et 6) telle qu’elle se déduit du courant expérimental correspondant par déconvolution des processus diffusionnels. |
Cette précision nous a permis d’élaborer le premier modèle entièrement cohérent de l’exocytose vésiculaire. Ce modèle démontre pour la première fois le rôle actif et déterminant des gels polyélectrolytes contenus par ces vésicules sur la cinétique de l’exocytose, alors que ces matrices polyélectrolytes n’étaient généralement perçues par les neurobiologistes que comme un moyen astucieux sélectionné par la nature afin d’encapsuler les cations catécholamines sous une forme très concentrée à l’intérieur des vésicules.
En effet nos résultats démontrent que la tendance bien connue (cf. Joanny-de Gennes) des gels polyélectrolytes à gonfler lors de l’échange d’un cation structurant (ici, catécholamine) par un monocation (ici, sodium ou proton de la solution extracellulaire) augmente la pression interne de la vésicule ce qui provoque un accroissement continu de la tension superficielle de la membrane vésiculaire. Ce phénomène persiste jusqu’au moment éventuel où l’énergie de tension superficielle du pore transmembranaire compense son énergie de bord (voir partie gauche de la figure 5).
| Figure 5 |
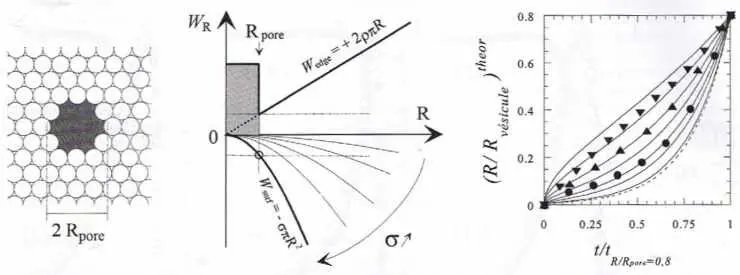 |
| Dynamique initiale de la fusion pendant la phase où la membrane reste sous tension. Gauche : schéma d’un pore transmembranaire et variation de ses deux composantes énergétiques (tension de bord, Wedge µ + R ; tension superficielle ; Wsurf µ ‑R2) en fonction de son rayon R. Au début de l’exocytose, la membrane n’est pas sous tension suffisante et le pore reste donc stable à son rayon Rpore imposé par son architecture biologique. Pendant la phase I (figure 2, émission via le pore transmembranaire), l’altération progressive de la matrice contrainte géométriquement par la membrane vésiculaire toujours intacte augmente la pression interne DP de la vésicule et par conséquent la tension superficielle (s µ DP) de sa membrane. Cela continue jusqu’au point de rupture du pore (courbe Wsurf indiquée en gras), ce qui détermine le début de la phase II de la figure 2. Tant que la zone équatoriale de la matrice n’est pas démasquée par l’expansion du pore, la pression interne continue à être transmise à la membrane qui reste donc sous tension. La vitesse d’expansion du pore est essentiellement alors contrôlée par la dissipation visqueuse de cette énergie de tension superficielle (en R2) puisque l’énergie de bord du pore est alors négligeable (énergie en R ; voir schéma). Cela provoque une expansion irréversible et rapide du pore (courbes de droite ; courbes pleines : famille de prévisions théoriques ; symboles : variations expérimentales pour trois événements vésiculaires différents). Ces processus ont lieu avec une cinétique initiale différente selon la fraction d’énergie de courbure Gaussienne non dissipée mécaniquement lors de la rupture explosive du pore, d’où la grande variabilité observée expérimentalement. |
Si ce point peut être atteint, le pore devient nécessairement instable et doit se rompre, ce qui provoque la fusion irréversible de la membrane de la vésicule avec celle de la cellule (début de la phase II, figure 2). Cette phase expose de plus en plus de surface de la matrice à la solution ce qui se traduit par une montée rapide du flux de neurotransmetteur (figures 3, 4). La fusion se poursuit alors irréversiblement jusqu’à ce que la matrice (en moyenne, encore pleine à 70–80 % de cations catécholamines) soit entièrement exposée à la surface de la cellule (phase III, figure 2). La matrice continue alors à se vider par diffusion sphérique inverse ce qui commande la cinétique de décroissance du flux observée sur les figures 3 et 4.
| Figure 6 |
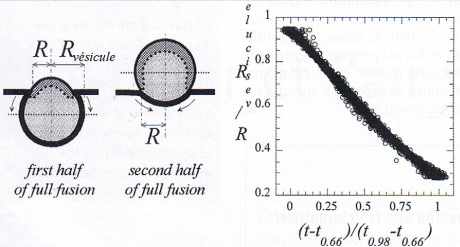 |
|
Dynamique la fusion lors de la phase finale de la fusion (passage de la phase II à la phase III de la figure 2). À gauche : différences entre les topologies des phases initiale et finale du processus de fusion suivant que la zone équatoriale de la vésicule a été ou non démasquée. À droite, variations expérimentales avec le temps du rayon de la zone torique de contact membrane – matrice après que la zone équatoriale de la matrice a été démasquée (schéma, dessin de droite). 596 événements individuels sont représentés sur cette figure. t0.66 : instant où les 2⁄3 de la surface de la vésicule sont démasqués ; t0.98 : instant où 98 % de la surface de la vésicule est démasquée (début de la phase III de la figure 2). |
La précision des courants mesurés par notre méthode de » synapse artificielle » permet d’exploiter quantitativement chacune de ces trois phases et de remonter au comportement dynamique de la membrane soumise à la tension créée par le gonflement du polyélectrolyte. En effet, tant que la membrane de la vésicule emprisonne encore la matrice polyélectrolyte (c’est-à-dire avant que son plan équatorial soit démasqué, voir schéma de gauche sur la figure 6), le gonflement de la matrice suite à son altération maintient la membrane sous tension, de sorte que l’énergie du » pore » est essentiellement d’origine superficielle.
Son expansion est alors contrôlée principalement par la dissipation visqueuse (cf. Brochard-de Gennes) de cette tension superficielle (en R2, figure 5). Le pore accroît ainsi son diamètre quasi exponentiellement (figure 5, droite). Cependant on observe une très forte variabilité due à celle de la cinétique initiale qui est fonction de la fraction de l’énergie de courbure Gaussienne non dissipée mécaniquement lors du microcataclysme lié à la rupture du pore initial (figure 5, droite).
Lorsque la zone diamétrale de la vésicule est démasquée par la membrane (voir schéma de la figure 6), le gonflement de la matrice polyélectrolyte n’exerce plus aucune pression sur la face interne de la membrane qui relaxe donc sa tension superficielle. Par conséquent, le transfert de la membrane vésiculaire dans la membrane cellulaire se fait avec une variation d’énergie de tension superficielle quasi nulle. Seule reste donc à prendre en compte l’énergie bien plus faible liée à la présence de la structure torique de la membrane au niveau de la jonction vésicule – membrane cellulaire (figure 6, dessin de droite du schéma). L’énergie de ce tore est équivalente à celle d’une énergie de bord, et par conséquent proportionnelle au périmètre interne du tore. Elle tend donc à se dissiper en diminuant son rayon. La dissipation visqueuse de l’énergie ainsi relâchée provoque donc une variation linéaire du rayon de la zone de jonction torique (figure 6).
Ces résultats démontrent que la dynamique de l’ensemble du processus de fusion est entièrement contrôlée par la dissipation visqueuse de l’énergie du pore en expansion dans une membrane initialement sous tension (figure 5 ; figure 6, dessin de gauche du schéma) puis relaxée dès que le plan diamétral de la vésicule est démasqué (figure 6, dessin de droite du schéma).
La matrice polyélectrolyte joue ainsi un rôle déterminant, en premier lieu en provoquant la rupture du pore initial (figure 5, gauche), puis en maintenant la membrane sous tension pendant la première phase de la fusion (figure 5 ; figure 6, schéma de gauche). À l’issue de la phase II (figure 2), la matrice polyélectrolyte n’a relâché que 20–30 % du neurotransmetteur qu’elle contenait initialement et se trouve complètement exposée au fluide extracellulaire. Le reste du neurotransmetteur est donc relâché ensuite par simple diffusion sphérique, ce qui explique la forme caractéristique des pics observés.
Conclusion
Outre le fait de proposer une première vision entièrement cohérente des processus exocytotiques vésiculaires et d’en dégager les grands moteurs physico-chimiques, ce modèle suggère aussi, et pour la première fois, que la taille des vésicules contenant le neurotransmetteur joue un rôle crucial sur le régime d’exocytose.
Ainsi, l’on prévoit que pour les petites vésicules, c’est-à-dire celles qui ne réussiront jamais à développer une pression interne (et donc une tension superficielle) suffisante pour contrebalancer l’énergie de bord du pore initial, l’on devra observer exclusivement une émission lente et à flux constant réglée uniquement par la taille du canal ionique initial. Ce dernier restera donc stable, maintenu par son architecture biologique lipidique ou protéique, et l’exocytose se déroulera uniquement via la phase I (figure 2).
Au contraire, les plus grosses vésicules arriveront à développer une pression interne suffisante avant que leur matrice ait pu relâcher la totalité de leur contenu cholinergique. L’émission lente via le canal ionique initial conduira ainsi inéluctablement à la rupture explosive du pore initial, et donc à une transition vers la phase II (figure 2) puis vers la phase III (figure 2).
Sur la base de ce modèle et des données quantitatives extraites de nos expériences sur les cellules chromaffines, et en faisant l’hypothèse chimiquement raisonnable que les paramètres énergétiques microscopiques varient peu d’un type de vésicule à l’autre, on peut fixer à 25–30 nm (soit 10–20 000 molécules) la taille limite des vésicules qui gouverne la mise en place de l’un ou l’autre de ces deux comportements exocytotiques.
Ainsi, les vésicules synaptiques du système nerveux central (25 nm environ, contenant de 10 000 à 20 000 molécules de catécholamines) correspondraient précisément à un système optimisé compte tenu de leur fonctionnalité. Ce seraient en effet les vésicules les plus chargées en neurotransmetteur tout en étant incapables d’aller jusqu’à la phase de fusion (exocytose via la phase I uniquement). Elles délivreraient donc un flux de neurotransmetteur constant, pouvant être interrompu par la fermeture de leur pore transmembranaire. Inversement, les plus grosses vésicules (glandes, mastocytes, etc.) seraient sélectionnées pour conduire irréversiblement à une fusion complète (sauf si bien entendu leur pore transmembranaire se ferme avant que la pression interne ait atteint la limite de rupture du pore). Cela permettrait en particulier de provoquer une exocytose massive et infiniment plus rapide que par le pore transmembranaire.
Ces prédictions semblent parfaitement cohérentes avec ce qui est connu biologiquement en ce qui concerne les tailles et les fonctionnalités de ces différents types de vésicules, pourtant, sans qu’aucun lien de causalité n’ait jamais été imaginé entre taille et fonctionnalité auparavant.
Enfin, cette vision suggère peut-être aussi une explication possible et originale du rôle néfaste des cations di- et trivalents sur l’exocytose. En effet, l’échange des cations catécholamines par des ions di- et trivalents intervenant en concurrence avec le sodium et les protons devrait correspondre à une capacité de gonflement plus réduite (peut-être même à une contraction du gel polyélectrolyte ; cf. de Gennes) de la matrice polyélectrolyte. Cela devrait d’une part diminuer ainsi la diffusivité des cations catécholamines et donc les flux d’émission du neurotransmetteur pendant la phase I, et d’autre part réduire la pression interne de la vésicule et partant, et pour le moins, amoindrir sa capacité à arriver à un état de fusion total.